|
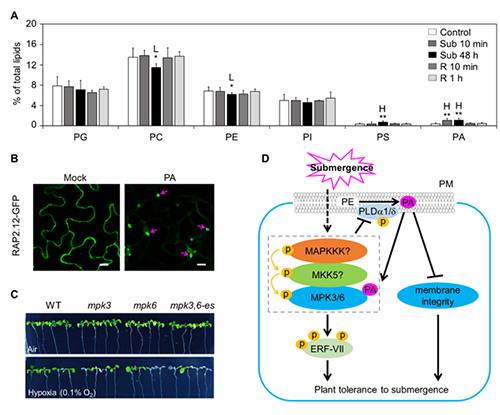
图1 磷脂酸PA调控植物低氧信号转导的分子机制,(A)脂质组学揭示水淹低氧处理显著诱导拟南芥中PA的积累;(B)体外PA脂质体处理可激活低氧核心转录因子RAP2.12-GFP融合蛋白由质膜向细胞核转移;(C)与PA互作的MPK3/MPK6激酶缺失突变体对低氧逆境表现出超敏感表型;(F)磷脂酸PA调控植物低氧信号转导的模式图
在国家自然科学基金项目(项目编号:31725004、31970298和32000215)等资助下,中山大学肖仕课题组在磷脂酸PA调控植物低氧信号转导的分子机制研究中取得进展。相关研究成果以“拟南芥磷脂酸调控MPK3和MPK6介导的低氧信号转导(Phosphatidic acid modulates MPK3- and MPK6-mediated hypoxia signaling in Arabidopsis)”为题,于2021年11月29日在线发表在《植物细胞》(The Plant Cell)上。论文链接:https://academic.oup.com/plcell/advance-article/doi/10.1093/plcell/koab289/6446039?searchresult=1。
低氧是影响植物生长发育与产量最常见的非生物胁迫之一。洪涝/水淹造成的淹没或积水降低了植物所处环境中的氧气浓度,使细胞处于缺氧状态,从而影响植物正常生理代谢和生长发育,导致作物减产甚至绝收,威胁农业安全。因此,研究植物对低氧胁迫的感知和信号转导机制,对于深入理解植物水淹适应性、保障洪涝灾害后作物稳产具有重要的科学和实践意义。目前,植物低氧响应的生理适应性机制已较为清晰,而对于植物低氧感知和信号转导的调控机理知之甚少。
研究表明,乙烯反应转录因子ERF-VII家族蛋白与膜相关ACBPs蛋白互作,锚定在质膜上。在水淹条件下,植物通过低氧诱导多不饱和acyl-CoA信号分子及依赖于类泛素化N-末端蛋白质降解机制,来实现对低氧胁迫的动态感知和应答,这些发现为植物脂信号分子参与调控低氧响应研究开拓了新思路。肖仕课题组利用脂质组学技术,系统分析了拟南芥在水淹前后和水淹恢复过程中的脂质含量,发现植物在水淹处理10分钟时PA含量快速积累,且在较长时间处理(48 h)仍维持较高水平。PLD?1和PLD?是P?产生的重要来源,其单突变体pld?1、pld?和双突变体pld?1pld?在低氧胁迫下呈现敏感表型。并且,PA与GST-MPK3和GST-MPK6重组蛋白存在体外互作,内源PA含量影响低氧下MPK3和MPK6蛋白激酶的磷酸化水平,暗示PA可能通过调控MPK3和MPK6磷酸化来影响植物低氧耐受性(图1)。
遗传学、生物化学和脂质组学证据揭示,PA激活的MPK3和MPK6依赖上游未知MAPKKK和MKK5,磷酸化植物低氧响应核心转录因子RAP2.12,调控其转录活性及低氧信号转导通路(图1)。同时,长期低氧导致PA的过量产生会造成活性氧(ROS)爆发、细胞死亡、及细胞膜完整性遭到破坏,不利于低氧去除后植物在复氧过程中的恢复生长。深入研究发现,MPK3和MPK6激酶通过直接与PLD?1和PLD?蛋白互作,磷酸化并负反馈调控其蛋白水平,从而维持低氧下合适的PA水平和细胞内稳态,保护植物在复氧过程中正常的生长发育(图1)。
该研究首次揭示了由PA-MPK3/6-RAP2.12模块介导植物低氧信号链的新机制,对完善植物低氧信号转导机理和作物抗涝遗传分子改良具有重要意义。