1988年,加州大学洛杉矶分校Michael Grunstein教授团队发现,真核细胞中的核小体不仅仅是供DNA缠绕的结构,它在调节基因表达上还有着重要作用1。此后,随着表观遗传学先驱David Allis教授在1996年对组蛋白尾部修饰酶的突破性发现2和2001年提出的Histone Code假说3,人们对染色质结构与基因转录调控之间关系的理解进入了崭新的一页。在一系列组蛋白新修饰被鉴定4的同时,人们迫切地想知道这些特异性修饰的性质和功能有何异同,又是如何调控基因表达的?例如,越来越多的证据表明,来自环境的代谢物会通过染色质的化学修饰实现基因转录调控5,但作为表观遗传调控的重要分子基础,人们对很多化学修饰,尤其是位点特异、双向可逆的动态修饰对基因表达调控的影响尚不清楚,亟需对这些多层次的表观遗传信息加以解析和关联。

论文截图
传统的生物学方法往往借助抗体的特异识别来原位研究蛋白质的翻译后修饰,而化学生物学技术能够实现活体环境下的高选择性和时空特异研究,具有独特的优势。在这一工作中,作者借助遗传密码子拓展策略,发展了一种具有单氨基酸位点分辨率的多组学技术Single-site-resolved multi-omics(SiTomics,图1)。通过将一系列带有赖氨酸酰化修饰的“光交联”非天然氨基酸,以位点特异的方式引入活细胞内的组蛋白当中,原位模拟内源修饰在基因组上的分布,并与蛋白质光交联和组学鉴定技术、基因组测序技术等相结合,他们系统性地开展了由位点特异的化学修饰介导的染色质相互作用蛋白质组与基因组“关联鉴定”。利用SiTomics技术,他们首先鉴定出H3K56位点在短链脂肪酸代谢物的刺激下会发生显著的酰化修饰,进而获得了受H3K56酰化修饰介导的相互作用蛋白质组和基因组,建立了二者之间的直接关联。他们的工作还揭示了超级增强子(super-enhancer)在细胞“代谢-修饰-基因转录”调控轴中发挥的重要作用。
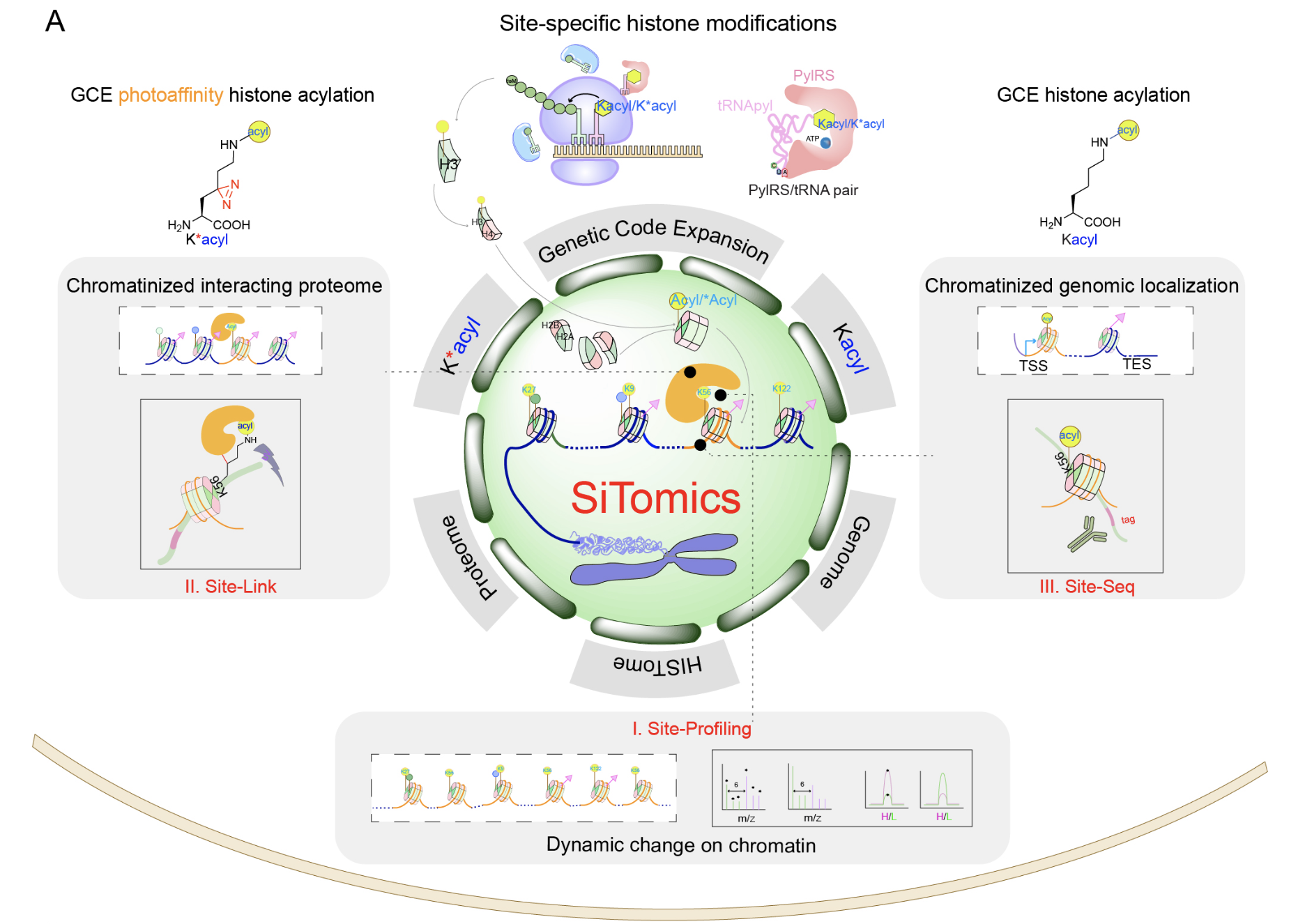
图1“关联解析”染色质动态修饰机制与功能的“单位点-多组学”技术-SiTomics(Single-site-resolved multi-omics)。I. Site-Profiling: 单位点修饰的动态变化情况的比较, II. Site-Link:通过遗传密码子拓展技术在组蛋白上引入K*acyl,利用光交联蛋白质组学鉴定单位点修饰的相互作用蛋白质组, III. Site-Seq:通过遗传密码子拓展技术在组蛋白上引入Kacyl,结合传统ChIP-seq技术研究具有修饰的核小体在基因组中所处的位置
“单位点分辨”的动态修饰定量鉴定
组蛋白修饰位点众多,不同位点对代谢物响应程度的定量比较,会因不同多肽对质谱响应的差异而难以实现。在之前的研究中,人们往往利用同位素标记的代谢物处理细胞并鉴定同位素标记的修饰肽段,但这样的方法会损失一些动态变化信息,尤其是对修饰量减少或者原有修饰经擦除后重新被修饰的情况,都难以提供丰度信息。利用SILAC技术标记赖氨酸和精氨酸,整合平行比较同样长度、修饰和电荷数的同一肽段的变化情况,可以更为精准地定量比较修饰的动态变化(Site-Profiling, I)。最终,通过加权分析,作者鉴定出H3K56位点在短链脂肪酸(如巴豆酸、β-羟基丁酸)处理下都显著地发生了相应的酰化修饰。
“非天然氨基酸”可于活细胞中“连接”表观遗传信息
如何在活细胞内实现染色质化学修饰的原位模拟?基于遗传密码子拓展的非天然氨基酸技术的优点正在于此。天然赖氨酸的光活性类似物-photolysine能够维持修饰状态与体内的赖氨酸修饰一致(图2),而遗传密码子拓展技术的生物“正交性”使得带有翻译后修饰的photolysine能够定点特异地引入组蛋白当中。借助这些“可遗传编码”的非天然氨基酸,带有特异修饰的组蛋白就可以通过其末端的富集标签,结合“光交联”蛋白组学技术(Site-Link, II)和传染色体免疫共沉淀-测序技术(Site-Seq, III),实现其相互作用蛋白质组及在染色体中定位的协同鉴定。
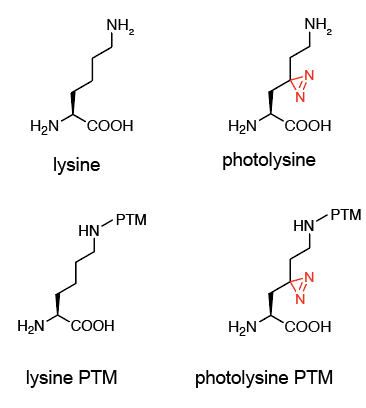
图2 Photolysine模拟lysine,最小化结构改变,同时便于维持PTM的结构基团稳定
那么,定点引入翻译后修饰的组蛋白是否能够稳定存在并被整合入染色体当中?被整合入染色体中的组蛋白修饰又去了什么样的位置?有没有干扰细胞内原始的修饰状态?和内源产生的修饰在基因组的分布又是否一致?这样得到的功能性发现又是否与实际状态一致呢?为了回答这些问题,作者设计、合成和开发了分别含有6种修饰(Kac, Kpr, Kbu, Kcr, Khib, Kbhb;K*ac, K*pr, K*bu, K*cr, K*hib, K*bhb)或1种对照(PABK与PABK*,通过TCO小分子原位decage出无修饰的K与K*)的共计14个“非天然氨基酸”探针(图3),均实现了组蛋白H3中相应赖氨酸修饰位点的定点插入。利用一系列表征手段,分别对带有特异修饰的组蛋白表达、分布位置、插入位点等进行了确证。同时,利用重标氨基酸探针确证了修饰基团在细胞中的稳定存在时间。这些详实的数据确证了带有定点翻译后修饰的组蛋白能够在活细胞内稳定存在并被有效整合入染色体当中。
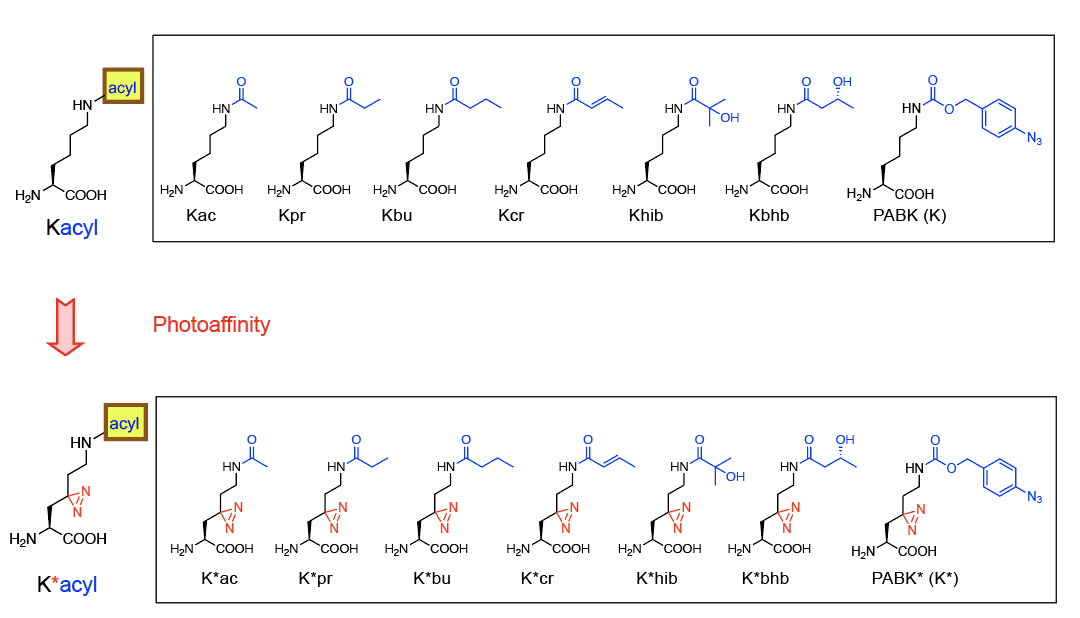
图3 本文所涵盖和发展的14个“非天然氨基酸”编码探针
互作蛋白质组与染色质分布的“协同解析”
作者将Site-Link技术应用于H3蛋白的K37, K56和K79等位点(H3K*56ac,H3K*56cr,H3K*56bhb,H3K*56PABK*,H3K*37cr,H3K*79cr),利用对这些带有光交联探针的组蛋白进行交联-富集和质谱鉴定,他们获得了不同位点/修饰的相互作用蛋白质组信息,发现不同“位点-修饰”组合的确具有独特的相互作用蛋白质组。
接下来,作者对巴豆酰化修饰进行了细致的研究(图4)。与H3K4me3一样,赖氨酸的巴豆酰化修饰(Kcr)被报道分布于基因组启动子区域。将SiTomics技术应用于H3K56cr的研究,作者确实捕获到一系列报道与H3K4me3相互作用的蛋白质,且随机选取H3K4me3在基因组中分布的位点能够验证H3K56cr也分布在这些位置。另外,他们发现H3K56cr具有一个非常特异的相互作用蛋白-GLYR1,与H3K56位的其他修饰基本都没有结合作用。鉴于之前的报道显示GLYR1分布在基因体(gene body)区域,作者推断GLYR1可能与巴豆酰化在gene body的相对高丰度分布有关。进一步利用“时间分辨”的光交联技术,他们发现在H3K56cr表达量稳定后,GLYR1与之的结合会逐步增加,这在一定程度上解释了H3K56cr是先被整合进入基因组中,再与GLYR1产生相互作用。在得到这些信息之后,作者推断细胞中可能存在着“正交”的“识别-装载”系统,能够协助这些携带有翻译后修饰的组蛋白整合进入染色体的相应位置,从而使这些组蛋白能够“重现”内源修饰在基因组上的分布,其获得的组学数据便是染色质化的蛋白质组和基因组学信息。为验证这一论断,作者随即对赖氨酸的β-羟基丁酰化修饰(Kbhb)进行了更为系统的研究。
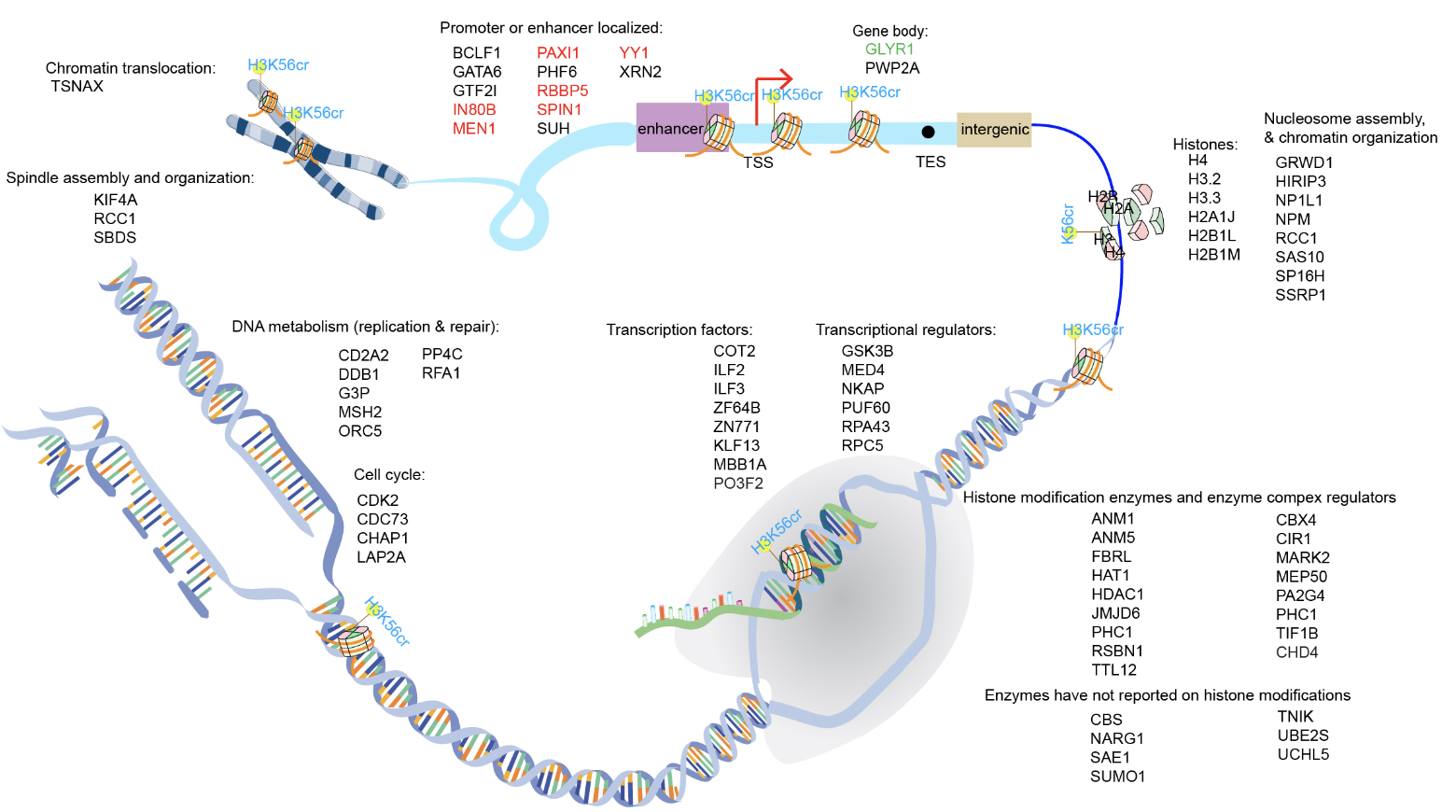
图4 SiTomics技术“协同解析”受H3K56cr介导的“染色质化(Chromatinized)”相互作用蛋白质组与基因组
β-羟基丁酰化是与“生酮饮食法”(Keto Diet)最为密切的一个化学修饰,但目前对其的研究报道甚少。将Site-Seq技术应用于β-羟基丁酰化,作者发现带有H3K56bhb修饰的组蛋白并非随机地分布在染色体中,而无修饰的H3K56则相对随机分布。H3K56bhb确实可以增加一些位置的染色体的开放性。进一步的Hi-C测序发现,单位点修饰的H3K56bhb便足以引起染色体三维结构的变化(图5)。
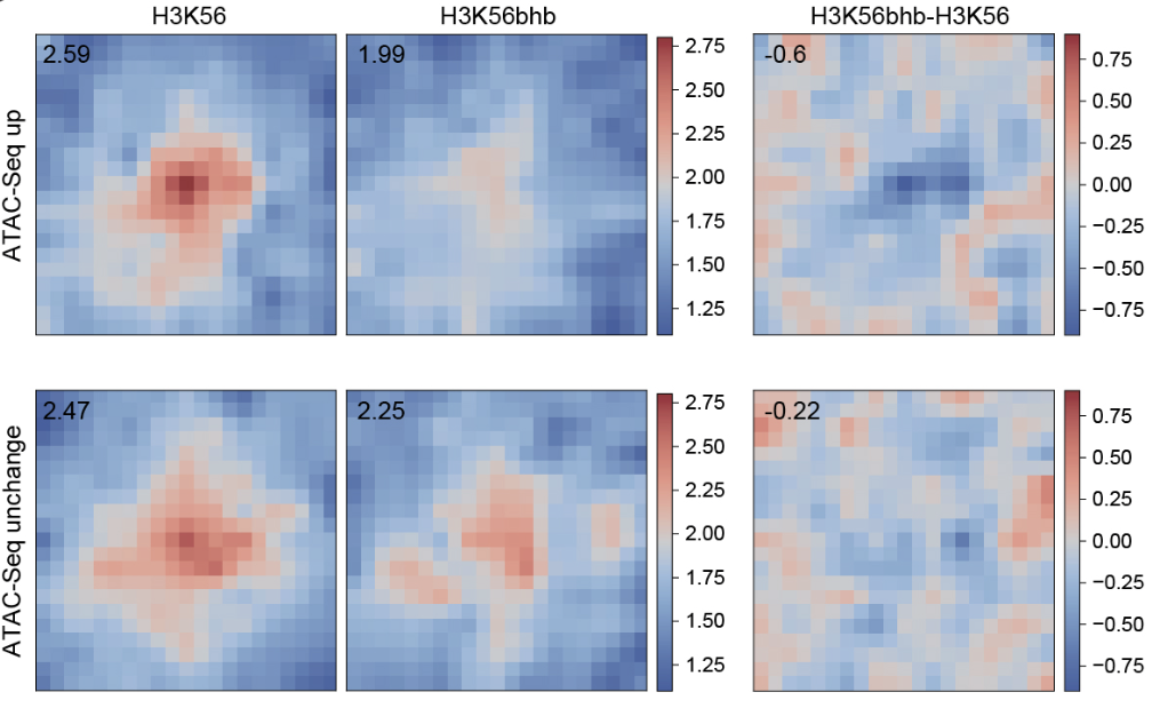
图5 H3K56bhb单位点修饰对三维基因组的影响
“单位点-多组学”技术揭示生理情况的染色体调控
在确认了工具对生理状态的模拟之后,作者接下来将其应用于酮体代谢下的染色体调控研究。他们首先通过免疫荧光发现酮体代谢下的β-羟基丁酰化修饰在细胞核中呈现特异的聚集样分布,这可能预示着细胞核中独特的调控作用。那么,如何快速寻找切入点,发现这样的生物学表型所对应的调控机制呢?他们所开发的“单位点-多组学”工具此时便有利于开启这样的探索。鉴于之前已经发现酮体代谢情况下,H3K56bhb修饰显著增加,则可利用SiTomics工具研究H3K56bhb的相互作用蛋白,并考察在基因组上是否存在特异分布。
的确,作者发现H3K56bhb与BRD4存在相互作用,同时,H3K56bhb在超级增强子中有显著分布(图6),而作为对照的H3K9bhb则几乎没有分布。作者因此推测可能酮体代谢情况下的染色体调控有超级增强子的参与。

图6 Site-Seq 分析H3K56bhb在基因组的特异分布
最后,通过利用Nabhb处理细胞、模拟酮体代谢下细胞内的染色质动态调控情况,作者发现,超级增强子的确发生了显著变化,而且与H3K56bhb的基因组整合位点相一致。此外,β-羟基丁酸处理下,受影响的超级增强子附近的基因其Pol II结合增加,而在H3K56bhb引入的细胞中,这些基因附近的Pol II结合也明显增加。进一步的RNA-seq分析证实这些发生变化的超级增强子可以帮助解释代谢情况下相关基因的高表达现象,从而在分子水平上实现了“代谢-修饰-调控”轴的贯穿(图7)。
图7 SiTomics解析代谢情况下H3K56bhb定点修饰对细胞超级增强子的影响
综上所述,SiTomics技术为“关联解析”染色质动态修饰的机制与功能、系统建立和解码表观遗传“关联信息”提供了多组学研究平台,并可适用于组蛋白之外的蛋白质和酰化修饰之外的化学修饰,为深入开展蛋白质动态修饰的机制研究、开发相应的化学干预策略提供了强大的工具平台。
陈鹏、季雄、北大-清华生命科学联合中心毕业生秦芳菲博士是本文的共同通讯作者;秦芳菲和生命科学学院李伯源博士是共同第一作者。文章返修过程正值北京疫情高峰期,多名同学贡献了自己宝贵的科研时间,展现了团结协作的风貌。
何川教授(HHMI、芝加哥大学)点评:
表观遗传研究近年来正在迅猛发展,DNA甲基化、染色质重塑、组蛋白的翻译后修饰、RNA可逆修饰7等都是表观遗传信息的重要载体(https://www.genome.gov/genetics-glossary/Epigenetics)。如何系统性地揭示环境与表型之间由表观遗传介导的动态关系,深入研究其分子机理,这就需要各种基因组学、成像和其它有效的方法的结合。表观遗传调控本质是生物大分子上化学动态修饰的问题。化学的方法来研究表观遗传应该有巨大前景。这篇文章提供了一个非常好的例子。其理论基础就是过去20多年广泛发展起来的生物正交反应体系。比如2022年的诺贝尔化学奖颁给了点击化学与生物正交反应,简单的炔基与叠氮基团的偶联反应,与生物体系兼容且正交,为很多生物学新发现提供了有效的工具。此次陈鹏团队与合作者的这个工作实现了利用遗传密码子拓展技术将组蛋白化学修饰定点“编码”到活细胞的染色质当中。他们系统分析了这些外源引入的化学修饰与生物体系内源修饰的正交性,同时利用光交联的化学手段研究相互作用蛋白质组,并结合传统生物学测序技术鉴定基因组信息,展示了这个技术的巨大前景。
1. 在技术发展方面,利用“遗传密码子拓展”策略,实现了活细胞内蛋白质翻译后修饰的“编码”研究
蛋白质的化学修饰通常都是“翻译后”事件,目前只能依赖特异识别某个修饰的抗体加以原位研究,尤其对于不同位点的相同修饰,甚至同一位点上化学结构相近的不同修饰,经常很难加以区分,更无法在活细胞内进行特异、原位的研究。通过使用化学合成的片段体外重组核小体可以对定点的组蛋白修饰开展结构和功能研究,但在活细胞内进行定点修饰研究仍然受限。普林斯顿大学的Tom Muir教授开发的intein技术虽然可以通过多肽/蛋白质的胞外递送在一定程度上实现活细胞内的研究,但是具有较多的局限性。遗传密码子拓展技术能够将这些带有修饰的氨基酸以“非天然氨基酸”的形式“编码”到活细胞的组蛋白当中,能够实现修饰类型和氨基酸位点的“精准编程”,为表观遗传修饰研究提供了广阔的天地。陈鹏团队发现修饰的氨基酸可以被“编码表达”到组蛋白的特定位点,而在染色质整体水平上这些“精准编码”的组蛋白能够维持稳定并在生理条件下被有效地整合进染色体当中。基于此,大量的组学数据也展示出染色体上这些结构相近的化学修饰的特异性。这一技术对未来针对特定修饰功能或两到多种修饰之间协同功能研究提供了一个普适平台。
2. 实现了“单个位点-特定修饰”的染色质相互作用蛋白质组-基因组“串联解析”
陈鹏与合作者发现组蛋白在单个氨基酸位点上的修饰变化就足以引发三维基因组的变化,为位点特异的组蛋白修饰调控高阶3D染色质结构和功能提供了有力的证据,也预示了这个技术的巨大前景。通过将SiTomics技术与ChIP-MS、ChIP-seq联用,他们以染色体上特定位点的化学修饰为单元,进行了蛋白质组、基因组的多层次“关联研究”,通过光交联同时可以在活体细胞里鉴定各种结合蛋白质,这些在以往的传统研究中是难以实现的。作者选择围绕显著受代谢影响的H3K56进行了各种功能研究,发现H3K56bhb在增强子上的调控作用,也展示了该技术在生物研究上的应用。
该工作有助于我们去理解受化学修饰调控的蛋白质组和基因组是如何相互影响,并最终影响基因转录的。我们希望这样的化学生物学工具越来越多,同时期待文中所提供的大量数据信息能够推动更为深入的生物学机制研究。